Plants are independent data set to test hypotheses of climate from marine record. They are spatially-fixed adapted to atmospheric and substrate conditions. They are tightly constrained by the climatic regime under which they grow.
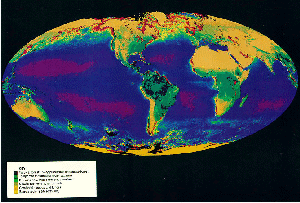
It can be seen from a WORLD VEGETATION MAP that there is a relationship between biomes and major climatic parameters, including radiation flux, temperature, precipitation.
Two approaches have been devised to use angiosperms in the analysis of past climates:
Depends upon level of taxon on which extrapolation is attempted. Where extant species are climatically restricted or climatic range well documented, approach may be better applied, especially in identical taxa. Applicable to Pleistocene but is highly age-dependent.
Bias introduced when utilizing only a single taxon as a basis for paleoenvironmental interpretations.
Physiological adaptions reflected in morphology, anatomy & chemistry. These are solutions to environmental stresses placed upon terrestrial organisms evolved early in plants, equilibration of temperature; water availability, nutrient supply; gas exchange and light capture. These solutions are taxon-independent.
Paleobotanical record is that of the solutions to environmental problems in a variety of spatial and temporal scales.
CUTICLE
Protection from animals & pathogens, mechanical injury, dust penetration, UV-radiation, temperature regulation and water loss. Biopolymers include Cutan/Cutin. Gas exchange important component, although earliest cuticularized plants do not possess exchange mechanisms. Gas exchange is facilitated by the presence of stomates.
Cuticle thickness is related to water stress. Thin cuticles characterize hydrophyllic plants; thick cuticles on water-stressed (high evapotranspiration, salinity, inefficient vascularization) plants. Sunken stomates (often overarched by papillae) effectively reduce water loss.
 Stomatal density related to environmental stress; monitor and record in C3 plants. Plants under high wind conditions
develop high stomatal density (but stomatal aperture is small); plants
subjected to higher pCO2 concentrations in the
atmosphere decrease the number of stomata per leaf area. high/low
CO2 content relationship. Stomatal size NOT a
good indicator, related to ploidy level of organism, but the
relationship between stomata and epidermal cells has been shown to be a
reliable
climate proxy.
Stomatal density related to environmental stress; monitor and record in C3 plants. Plants under high wind conditions
develop high stomatal density (but stomatal aperture is small); plants
subjected to higher pCO2 concentrations in the
atmosphere decrease the number of stomata per leaf area. high/low
CO2 content relationship. Stomatal size NOT a
good indicator, related to ploidy level of organism, but the
relationship between stomata and epidermal cells has been shown to be a
reliable
climate proxy.
Stomatal Index = Stomatal Density (#/sq area)/Stomatal & Epidermal Cell Density
LEAF FORM
Size - Chaney (1947) subdivided leaves based upon their overall dimensions: greater or smaller than 10 cm; large vs small surface areas. Large leaves indicative of non-limiting water availability during growth (although light flux, temperature and nutrients play a role).
Drip Tips - Attenuated apices common in rainforests assisting in shedding of water (and retardation of fungal and epiphyllous colonization). Presence/Absence
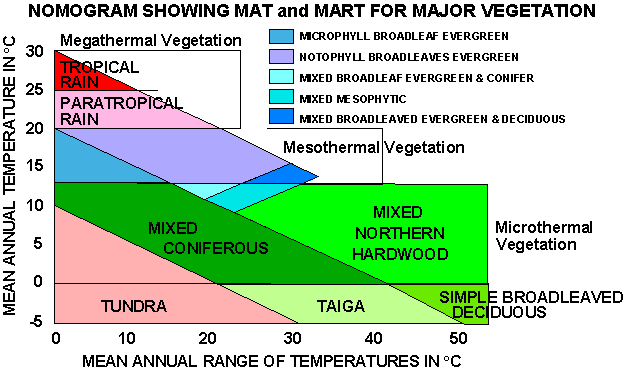
Leaf Margins - MAT has been correlated with dicot leaf margin characteristics (Wolfe, 1979). Ratio of entire margined leaves to those with serrations (teeth) correlates directly with MAT. Differences noted between northern and southern hemisphere floras (latter with many evergreen taxa). Assemblages must be evaluated taphonomically before utilization of this technique. Recently, Wilf (1997) has demonstrated that percentage of entire leaf margined leaves is directly correlatable with MAT.
Abscission - Evergreen (continuous production and abscission) vs deciduous (synchronous and temporally discrete) taxa is not easy to distinguish in fossil record. Factors include light intensity, day length, temperature change, nutrient flux, water stress, physical damage, infection all affect abscission. Synchroneity of abscission related to seasonal fluctuations in light, temperature and water.
Limits of solar energy input can be deduced from autochthonous (in situ) stump grounds in which wood anatomy is preserved. Amount of wood produced annually by an are of forest sets a lower limit to annual solar energy input. (Creber and Francis, 1987)
Assmann (1970) devised a method in which he calculates the cross-section area of the trunk added by each successive growth ring (extant wood). This is directly proportional to the amount of wood produced by the whole trunk during the year. Dependent not on ring widths alone but influenced by the diameters of the cross sections of trunks on which they accrue. Using incremental plots against years, Hamilton & Christie (1971) have demonstrated that annual productivity can be characterized as high (5 mm/yr), medium (3 mm/yr) and low (1 mm/yr). In fossil wood, the radius of the tree can be estimated from the curvature of the rings (taphonomic bias must be considered).
INTERPRETIVE DENDROCHRONOLOGY
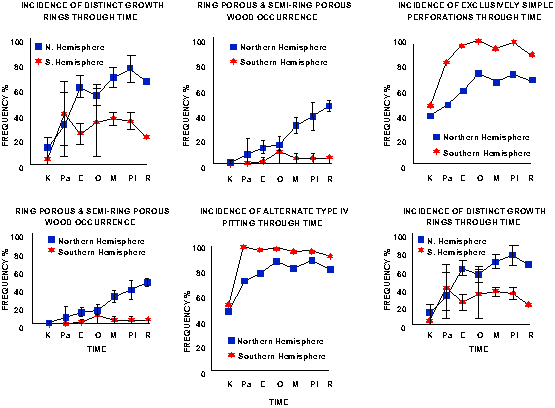
Deductions of climate can not be made on growth rings alone. Variation in species and growth sites, successive growth rings differ. Hence, the use of:
Mean sensitivity coefficient = Difference in consecutive ring pair width/Average ring width of sequence
| 0.3 | no yearly variation |
| >0.3-0.6 | water limited & variable |
| >0.6 | extremely restricted water supply |
Absence of growth rings under humid equatorial climate is common but not the rule. For example, well defined rings occur in:
In the Amazon, 22% of trees display poorly defined rings, and figures for angiosperms that do not produce growth rings occur include Amazon (43%), India (75%), and south Florida (42%).
Caveats to using coniferous wood:
Caveats to angiosperm wood:
Common in fossil record, but often overlooked. Roots have aeration systems that include aerenchymatous tissues. These are found within roots that grow under waterlogged conditions.
Root cuticle and Endodermal features - Regulation of water and nutrient uptake; their presence usually indicates plants growing in dry areas or mires periodically dried. Thickening of the endodermis is reported to play a role in ion selectivity and the development of hydrostatic pressure in the root system. Such features occur in unrelated fossil groups.
Mycorrhizal associations - confer greater drought resistance an nutrient access. Confirmed in the fossil record Triassic peat.

