CRETACEOUS ANGIOSPERM DIVERSITY
Several authors have evaluated the diversification of flowering plants from their initial appearance in the fossil record.
These patterns have been based either on megascopic or palynological remains. Lidgard and Crane evaluated absolute
species richness of leaf data (the number of taxa reported in the literature) for angiosperms, conifers, pteridophytes
(ferns, lycopophytes, sphenophytes), cycadophytes (cycads, cycadeoids, and pteridosperms with simple pinnate leaves),
ginkgoes (Ginkgo and Czekanowskiales), and other seed plants. Beginning about 90 MY ago (mid-Albian), there is a
widespread dominance in megafloral assemblages of the Northern Hemisphere. As can seen from their figures (below),
there appears to be competitive displacement of cycadophytes and pteridophytes.
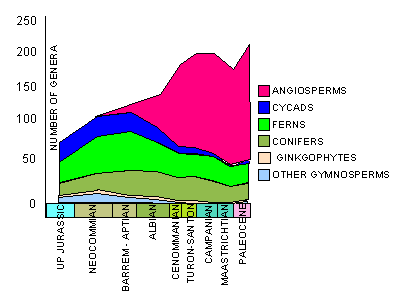
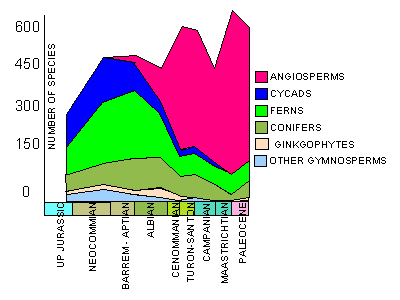
Between 115-90 MY, there was within floral diversification of the angiosperms, with essentially no response or
reaction within the coniferophytes. The rise to dominance with increased diversity seen in megafloral assemblages is
not paralleled coevally with the palynological record. Based on palynological data, angiosperm dominance is achieved
in the Turonian. Diversification occurred at a rapid rate, following a kinetic growth model (S-shaped growth curve).
Cycadophytes begin a decline in diversity before angiosperms become dominant, and the opening of this ecological
niche may have provided the opportunity for angiosperms to diversify.
Patterns of Diversification
- First angiosperms appear in low numbers in the Barremian. Their early history is confined to stream-margin habitats
at low latitudes (equatorial) and were secondary in importance (abundance and diversity) to ferns, cycadophytes,
and conifers. Several of these early angiosperms are "paleoherbs."
- Large trees are known for the first time in the early-middle Albian. Mid-Albian flowering plants occur in higher
middle latitudes and were locally dominant. Diversity equals that of ferns and other groups.
- Late Albian-Middle Cenomanian landscapes had angiosperms in environments spanning the tropics to the poles
inhabiting stream-margin and overbank (floodplain) facies. At this time, nearly all major physiognomic foliage types
can be found. There also appear to be strong latitudinal gradients in leaf types established by the Mid-Cenomanian.
- Coastal Plain floras reflect warm, equitable climates with a poleward displacement of major isotherms
- Megafloral assemblages from below 60oN consist of
broadleaf evergreens, with cold-sensitive life-forms
distributed I a pattern comparable with these plants.
- Arctic megafloras are dominated by broadleaved deciduous forests, the distribution of which was probably
controlled by winter darkness and possibly freezing temperatures.
- Cenomanian to Coniacian assemblages show a rich and diverse conifer assemblage in which there is evidence for
increasing diversity of angiosperms.
- Palms first appear in the Santonian in eastern North America and extend their range into higher middle
latitudes and other monocots have been documented.
- Dicotyledons were the most abundant element with five subclasses known: Magnolias, Hamamelids (e.g.,
sycamore), Roses, Buttercups, and Dilleniids.
- There is evidence for the first angiosperm halophytes, adapted to brackish-water conditions; true mangrove
swamps dominated by angiosperms may have originated no earlier.
- Prior to the postulated appearance of closed-canopy angiosperm forests, there is evidence to indicate that
megathermal rainforest existed in the mid-continent.
- Campanian to Maastrichtian leaf assemblages from the southern Rocky Mountains appear to be similar to those that
preceeded; those from south of the Arctic circle reflect generally subhumid conditions.
Cretaceous Vegetational types
Floral provinces of the Cretaceous are primarily based on
palynological suites, and during the Barremian-Cenomanian Brenner
has recognized 4 floral provinces:
- Northern Laurasian Province - temperate, humid, now
above 60oN; Bisaccate pollen of Pinaceae, NO Classopollis
(Cheirolepsis) or Ephedra-type pollen; monosulcate pollen of the
cycadophytes is rare. Fern pollen is abundant, but at a reduced
diversity than in lower latitudes.
- Southern Laurasian - warm, temperate to
subtropical. High diversity in the middle latitudes with
abundant and diverse ferns; coniferous pollen of Cheirolepidaceae
(Classopollis) dominating the spectrum (60-80%).
- Northern Gondwana - tropical to semi-arid with
bizarre forms of pollen; low fern spore diversity and a paucity
of Schizaceous ferns. NO Bisaccate Pollen of Pinaceae,
araucarian and podocarp pollen; Cycadophyte and Ephedraceae
pollen.
- Southern Gondwana - warm temperate to subtropical
climate. Podocarp bisaccate pollen abundant, pteridophyte
diversity.
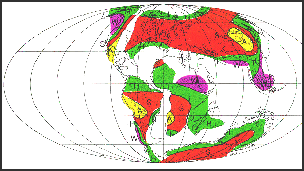 |
Predicted mean annual rainfall for the Cenomanian (from Parrish et al.,
1982). Wet=Purple; Humid=Green; Semi-arid=Orange; Arid=Yellow.
In the Late Cretaceous of North America, three vegetational types have
been recognized:
- Eastern North America and southern Western Interior supported subhumid, megathermal, broadleaved evergreen
woodlands. Conifers dominated with angiosperms, although comprising up to 80% of the assemblages, mainly
restricted to the understory.
- Northern Western Interior and Pacific Coast supported subhumid, broad-leaved evergreen forest living under
seasonally wet conditions. Higher diversity of vines, some species with drip tips. Probable deciduous trees have a
higher diversity. More angiosperms are known from Maastrichtian floras than Cenomanian to Campanian
assemblages. Cycadophytes and Gingophytes remain common in these sites.
- Arctic regions supported polar broad-leaved deciduous forests, which included both deciduous gymnosperms and
angiosperms. The southern part existed under cool mesothermal conditions; the northern part existed under
microthermal conditions.
Vegetational Response at the K/T Boundary
Takhtajan has subidvided the floras of the Late
Cretaceous into three paleobiogeographic zones within the Northern
Hemisphere:
- The Boreal realm is dominated by mesophyllous deciduous
trees and shrubs and possessing a high proportion of ginkgo,
conifers and ferns. This belt extends through North America, the
Arctic, Greenland, North-Central-Northeast Europe, Kazakhstan,
Siberia, etc.
- The Tethyan realm is characterized by a subtropical climate
with evergreen shrubs with an admixture of deciduous taxa. Most
leaves are narrow in form and entire (leaf physiogamy is a valid
indicator of climate, Wolfe 1979). The Southern US, England,
South Europe...Central Asia and China occupy this floristic
realm.
- The Paleotropical realm is preserved in Nigeria,
Egypt...Iran and the plants are similar to Malaysian-type
tropical vegetation in addition to mangrove habitats.
Widespread disruption occurred in parts of the northern hemisphere ecosystem. Palynological and megafloral evidence
indicates that there was a mass-kill event followed by a successional pattern of recovery. Ecological disruption is
evidenced by the relative abundance of characteristic pollen and spores, with a change from angiosperm-dominated
assemblages just below the K/T boundary to fern-dominated assemblages above. Above the K/T boundary, angiosperm
pollen markedly declines in abundance, with the relative abundance of fern spores rising to between 70-100%. This has
been termed the "fern spike." Studies of dispersed cuticles corroborate the palynological data set.
Northern vegetation shows less disruption at the K/T boundary than do floras from Eastern North America and the
southern Western Interior. This may be due to the development of more efficient dormancy mechanisms in these polar
forests. Plant groups in which dormancy mechanisms are not well developed appear not to have survived the K/T
boundary event successfully.
Two features of the climate/vegetational environment were significantly altered:
- precipitation levels and degree of canopy closure
- latitudinal juxtaposition of vegetational types, differing dramatically from the Campanian-Maastrichtian.
©Copyright 1997 by Robert A. Gastaldo. All rights reserved.
No part of or transmitted in any form or by any means, electronic or
mechanical, including information storage and retrieval system, without
permission from the author.

